大脑“合租区”的隐秘差异:单个神经元分辨率下解码性别共享神经系统的两性二态性
2025-08-08
前言:
近十年来,神经系统性别差异发展的分子机制研究受到人们的关注,在一些神经系统疾病的流行和进展中这种性别差异尤为明显,如神经分裂症和帕金森病等。在动物身上,不同性别驱动行为的独特回路又被称为“两性二态”,例如小鼠的移情行为回路,果蝇的攻击行为回路,以及蠕虫的伤害和机械感觉行为回路等。性别特异性行为通常归因于神经元连接和分子组成的差异,但基因性别如何在单个神经元水平上塑造神经系统的分子结构仍不清楚。由于哺乳动物系统中的激素调节可能会掩盖性别身份对单个神经元水平上不同分子特征的直接影响。因此,在今天这篇文献中,小编将为您介绍本文的研究者们是如何在模式动物上剖析单细胞分辨率下的两性二态神经元回路,揭示不同性别驱动行为中特定基因产生的贡献。
线虫秀丽隐杆线虫(C. elegans)的每个体细胞的遗传性别都是细胞自主决定:性染色体与常染色体的比例控制转录级联反应,从而导致雌雄同体特异性或雄性特异性基因的表达。早期也有研究者做过线虫秀丽隐杆线虫(C. elegans)的scRNA-seq,但是只关注了雌雄同体,对雄性神经元分子特征和两性分子特征差异的了解存在局限性。为了更广泛地了解秀丽隐杆线虫两性神经系统的分子结构,研究者们对雄性和雌雄同体的性别共享神经元类型进行了scRNA-seq转录组分析。
结果
1. 成年雌雄同体和雄性神经系统的scRNA序列分析
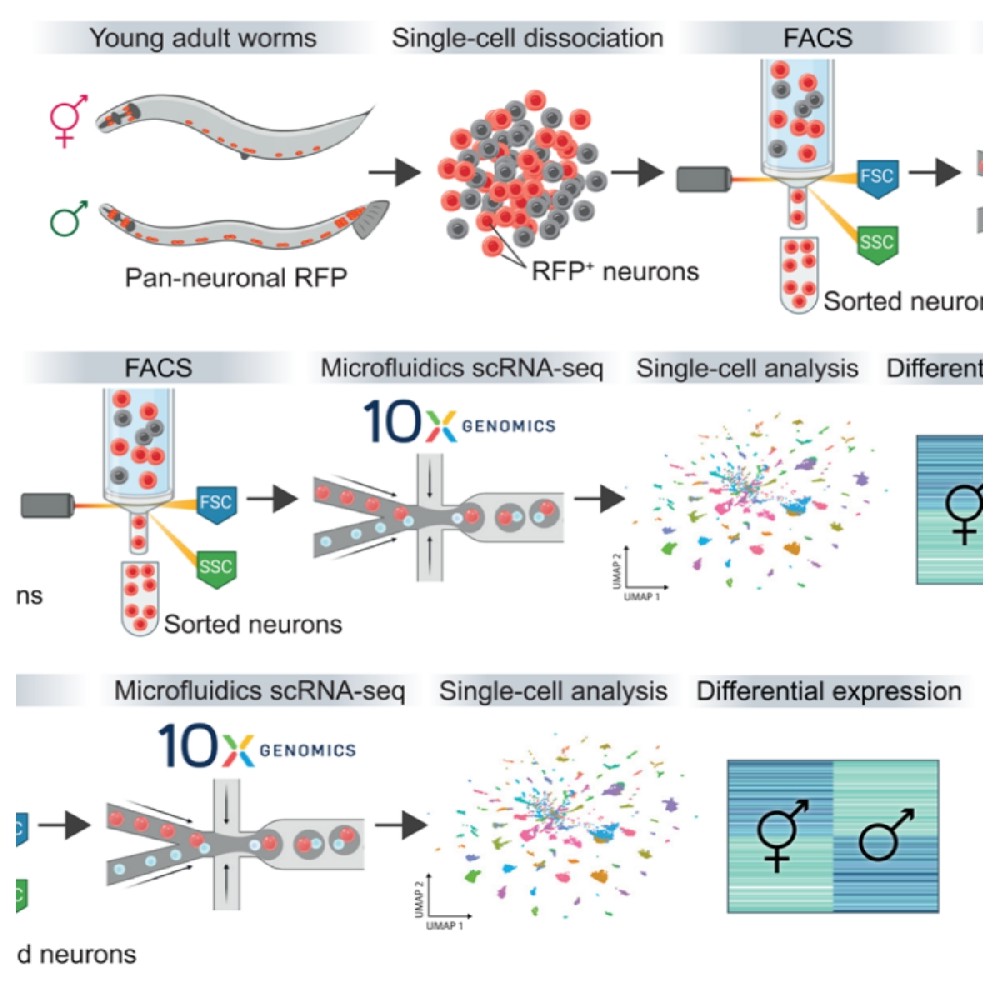
首先,研究者们做了大规模分离高纯度雄性的实验,产生纯雄性种群(XO动物)。接着,他们将年轻的成年动物组织分离成单细胞悬液、荧光分选(FACS)(图1A)、过滤出神经元,最终获得了雌雄同体的13015个单细胞神经元转录组和雄性的19454个单细胞神经元转录组数据,并进行聚类分析。
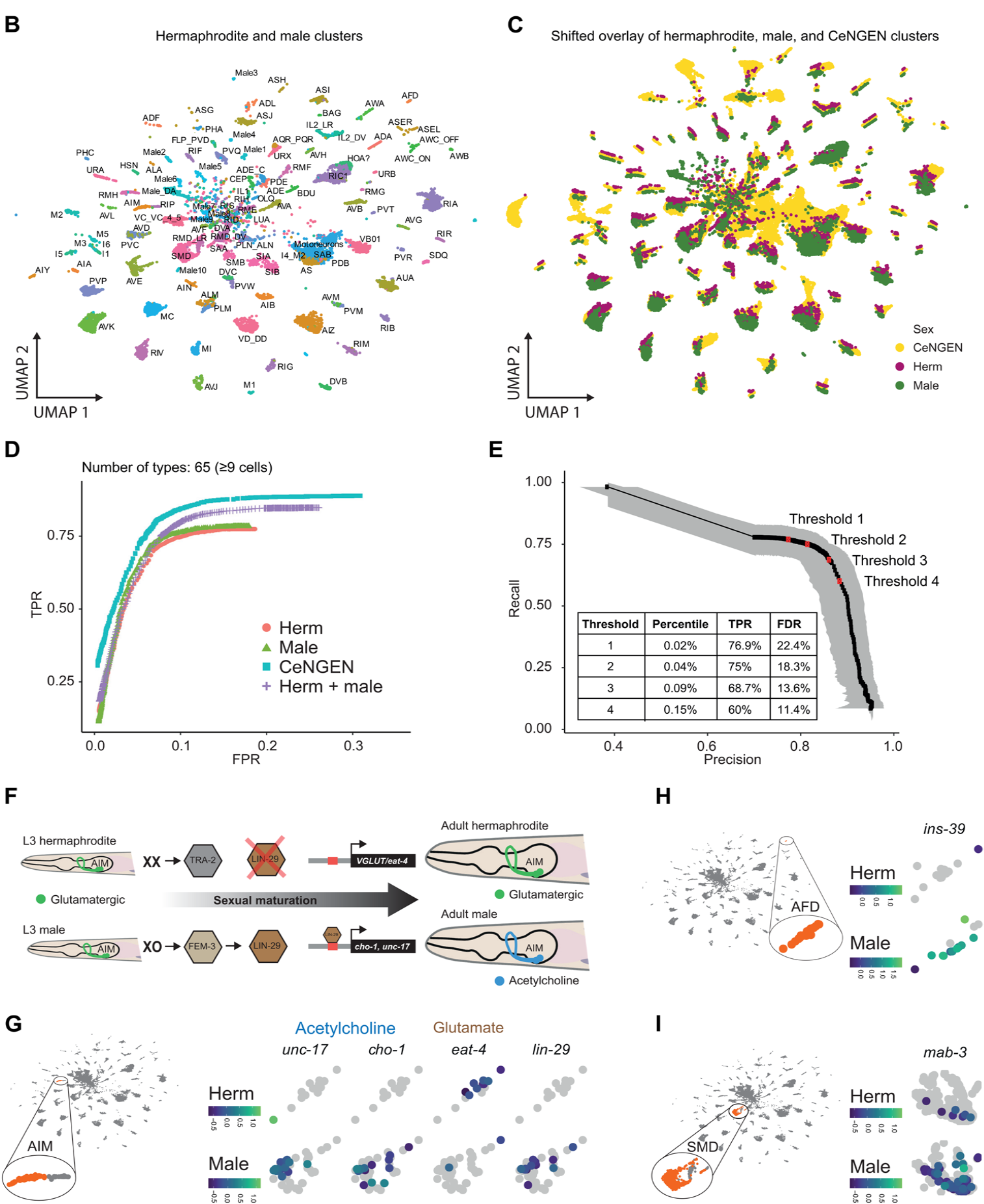
Fig1 秀丽隐杆线虫两性scRNA序列分析。
2. 神经系统的单细胞图谱捕获二态基因表达
对于基因表达分析,研究者们通过计算每个簇中表达一个基因的细胞的比例、平均表达和标记,设定一定的表达阈值,与细胞类型标记分析相结合,最终他们得到了能描述具有已知性别偏倚表达的基因表达谱。包含在雄性细胞群中、雌雄同体细胞群特异性表达的一些基因。如LIN-29、胆碱乙酰转移酶cho-1/CHOT和囊泡乙酰胆碱(ACh)转运蛋白unc-17/VACHT(图1F)、囊泡谷氨酸转运蛋白eat-4/VGLUT(图1G)、ins-39、mab-3等(图1,H和I)。
3. 性别共享神经元簇的比较分析揭示了广泛和多样的分子二态性
接下来,为了揭示性别共享神经系统的转录组谱在雄性和雌雄同体之间的差异程度,研究者们根据DEGs的数量对性别共享神经簇进行排序和过滤(详见原文材料和方法)。最终,分析捕获了已知的具有二态基因表达谱的性别共享神经元,包括PHC、AFD、AIM和AVG(图2A)。同时还揭示了许多未被归类为二态的性别共享神经元,如6个trn、BDU、PVR中间神经元等(图2A)。
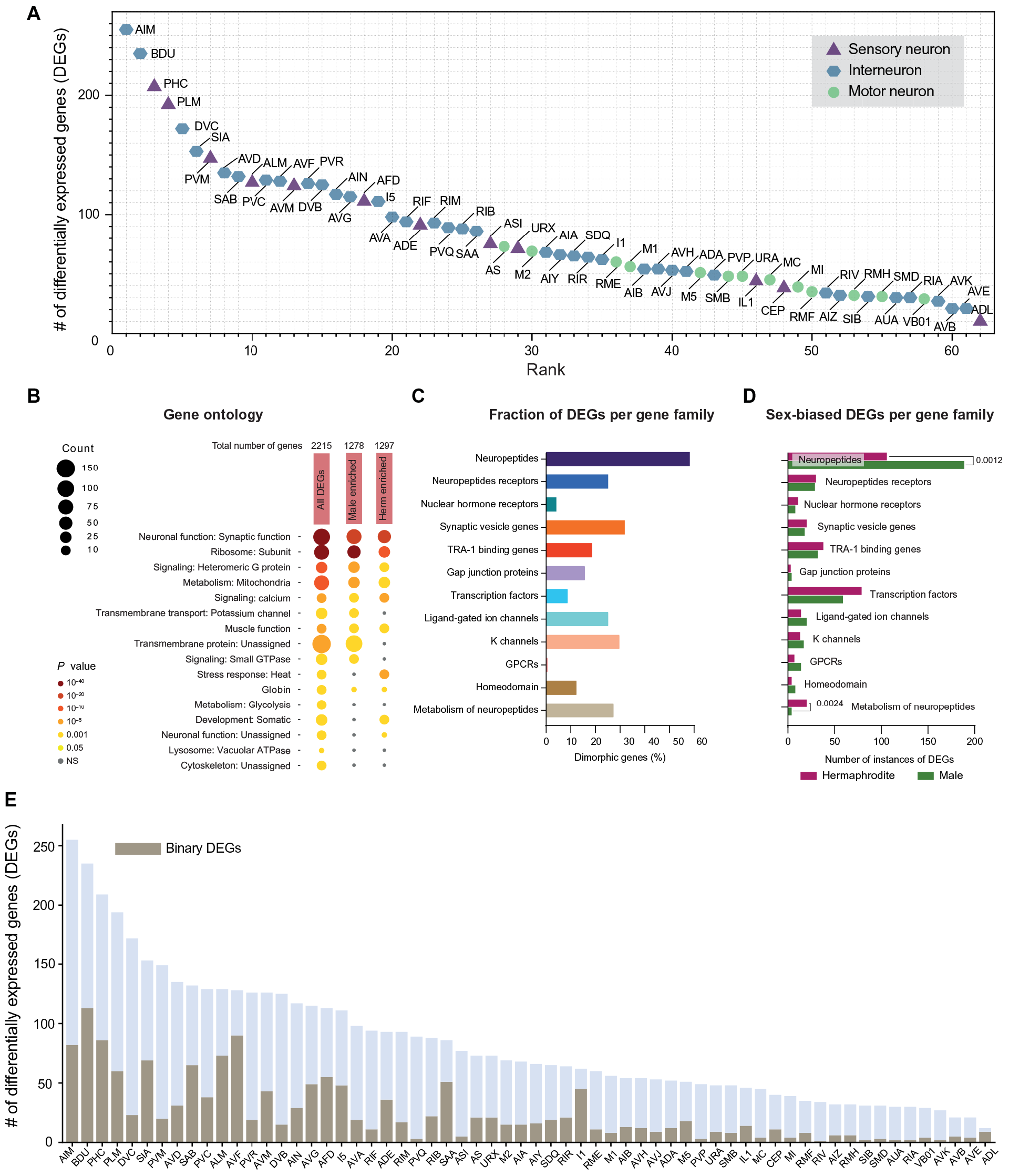
Fig2 性别共享神经元的比较分析揭示了广泛的分子二态性。
此外,排序分析共发现了所有性别共享的聚类中存在2215个DEGs,GO富集分析显示,与神经元功能相关的占比很大(图2B)。其中,神经肽、神经肽受体和神经肽代谢相关基因的过度表达,神经肽和神经肽代谢相关基因的男性表达明显存在偏倚(58.5%;157个预测的神经肽基因中有92个是DEGs基因)(图2、C和D、3A)。
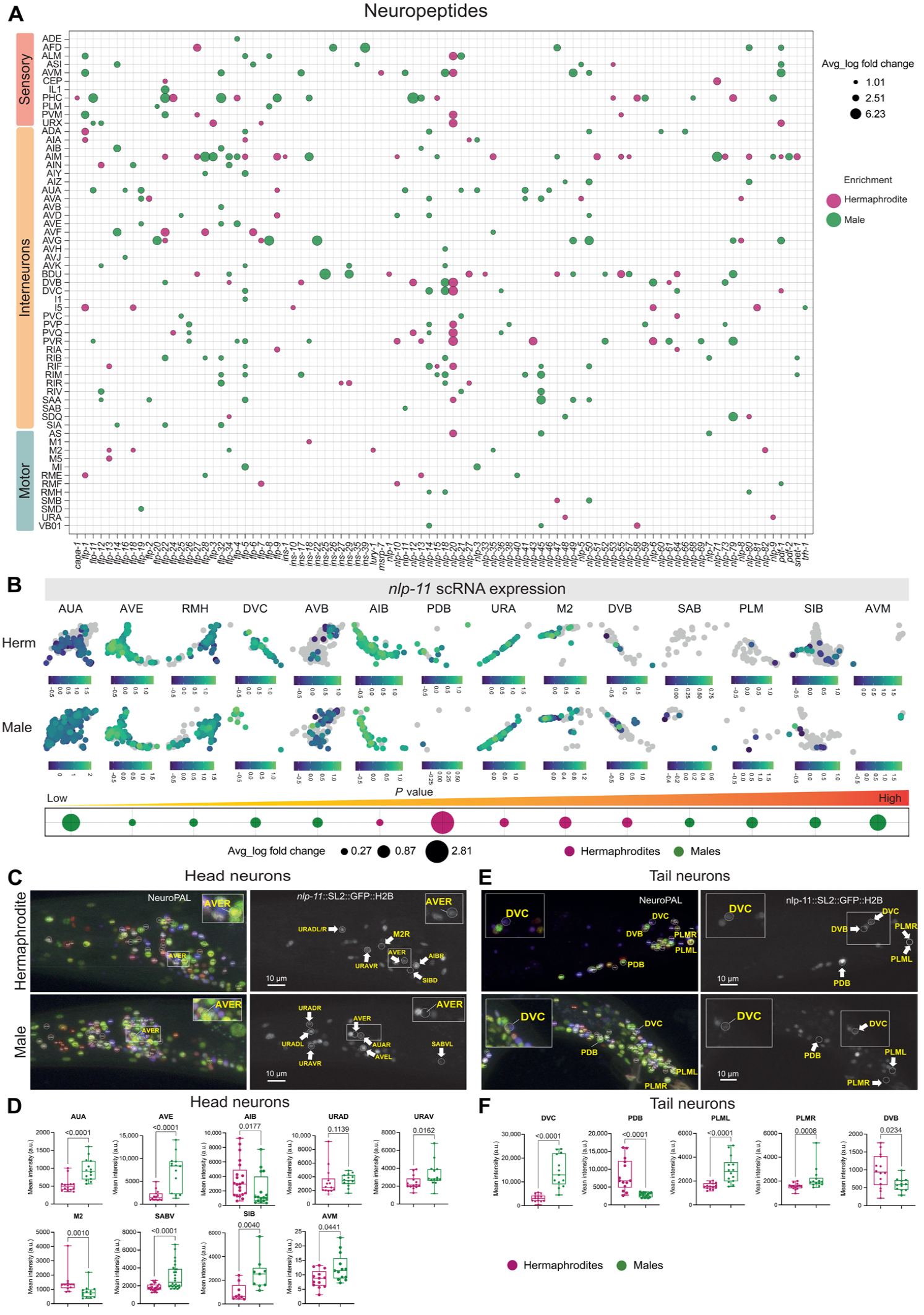
Fig3 神经肽在神经系统中的两性二态性。
研究者们在两个广泛表达的神经肽基因nlp-11和flp-20的表达模式中进行了性别二态性的预测和验证,发现,nlp-11在16个预测的性别共享神经元中的12个以及几个雄性特异性神经元中显示二态表达(图3,B至F)。flip -20在四个性别共享神经元和雄性特异性CEMs中表现出性别二态表达。
4. 多种神经元基因家族在两性之间的表达存在差异
除了神经肽基因家族外,其他基因家族也表现出神经元功能上性别差异(图2)。很大一部分突触相关基因和离子通道存在差异表达(图2C)。
5. 神经递质基因表达的性别差异在不改变身份的情况下塑造神经元输出
神经元末端身份由特定效应基因电池的表达来定义,而这些效应基因电池由被定义为“末端选择器”的tf直接激活和维持。研究者们比较了已知的末端选择器的表达,发现它们表达模式大多相似,只有少数例外。研究者们也检测了雌雄同体和雄性共享神经元簇中神经递质相关基因的表达(图2A和4A所示为相同的神经元簇,有些被排除在外,参见材料和方法)。虽然已知的AIM神经递质开关在数据集中被完全捕获(图1、F、G和4A),但他们没有观察到任何其他类似的神经递质身份开关(图4A)。然而,在检查这些神经递质相关基因的差异表达时,他们发现其中一些基因,主要是胆碱能相关和谷氨酸能相关基因,在几个性别共享的集群中表现出偏倚表达(图4B),这表明性别二态性可能在神经递质释放和合成方面更为突出,而不是在身份上。
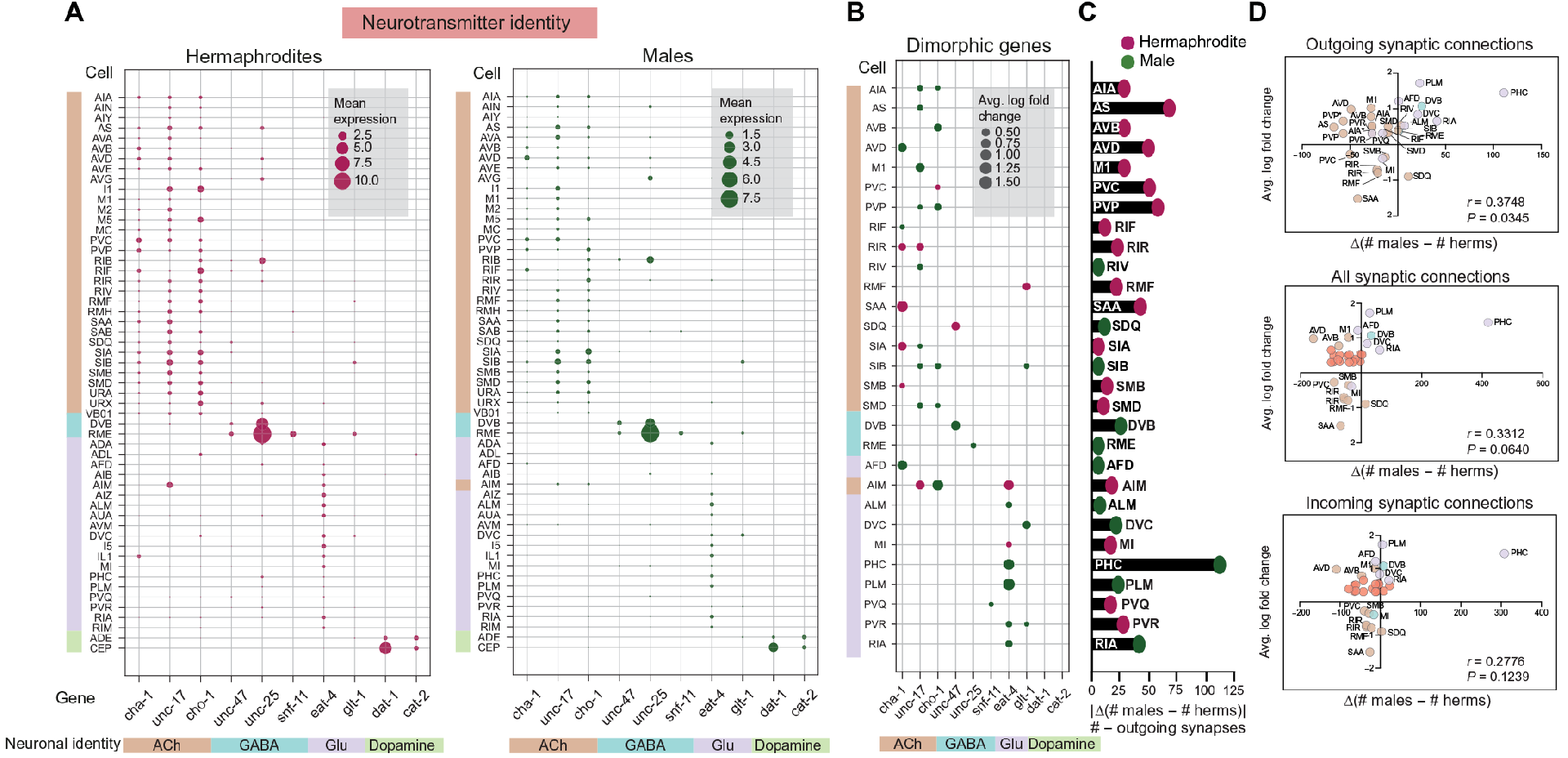
Fig. 4 神经递质身份在性别共享的神经元簇中保持不变。
基于这一点,他们做了二态神经递质相关基因表达与二态突触连通性之间的关系探讨。他们假设,在一个性别中表现出更高的神经递质相关基因表达的神经元可能也有更多的外向连接。为了验证,他们统计分析了两性之间在输出、输入和所有突触连接(化学连接)上的连通性差异(图4C)与图4B所示的神经递质相关基因的平均对数倍变化。值得注意的是,Pearson相关系数仅在外向连接中显著,而在传入或所有突触连接中不显著(图4D)。例如,在性别共享的神经元SIA、SAA、RIR和AFD中观察到乙酰转移酶cha-1的性别特异性富集,这与这些神经元在各自性别中的连接增加一致。相反,泡状谷氨酸转运体-4的性别特异性富集与神经元RIA、PLM、PHC、ALM和AIM在各自性别中的更大连接相对应(图4、B和C)。这说明神经元的神经递质释放与其外向连接是相关的。
总的来说,虽然神经递质身份在性别共享的神经元中很大程度上得到了保留,但神经递质相关基因的性别二态表达与外向突触连接的差异有关,这种机制表明功能二态可能在不改变核心神经元身份的情况下出现。
6. PLM神经元的性别二态基因表达和功能是性别特异性机械感觉反应的基础
接下来,研究者们在具有大量DEGs的性别共享神经元簇探索了对轻柔触摸的机械感觉反应所必需的所有TRNs: ALM、AVM、PVM和PLM(图2A),其中特别关注PLM,它不仅介导后部轻柔触摸反应,也是TRN中性别最二态的(图2A和5A)。
研究者们通过转录和翻译报告(图5,D至G)以及单分子荧光原位杂交(smFISH)(图5H)验证了多个神经元相关基因的差异表达(图5,B和C)。发现雌雄同体对后触的反应降低,但对前触的反应消失(图5I)。因此,他们推测,这种后轻触行为二态性可能源于PLM神经元活动的性别差异。当测量PLM基础活性的钙痕迹时,他们观察到在频率高达2 Hz的雄性中较高的活性(图5,J至M),这些实验结果进一步支持PLM在性别上的二态作用。
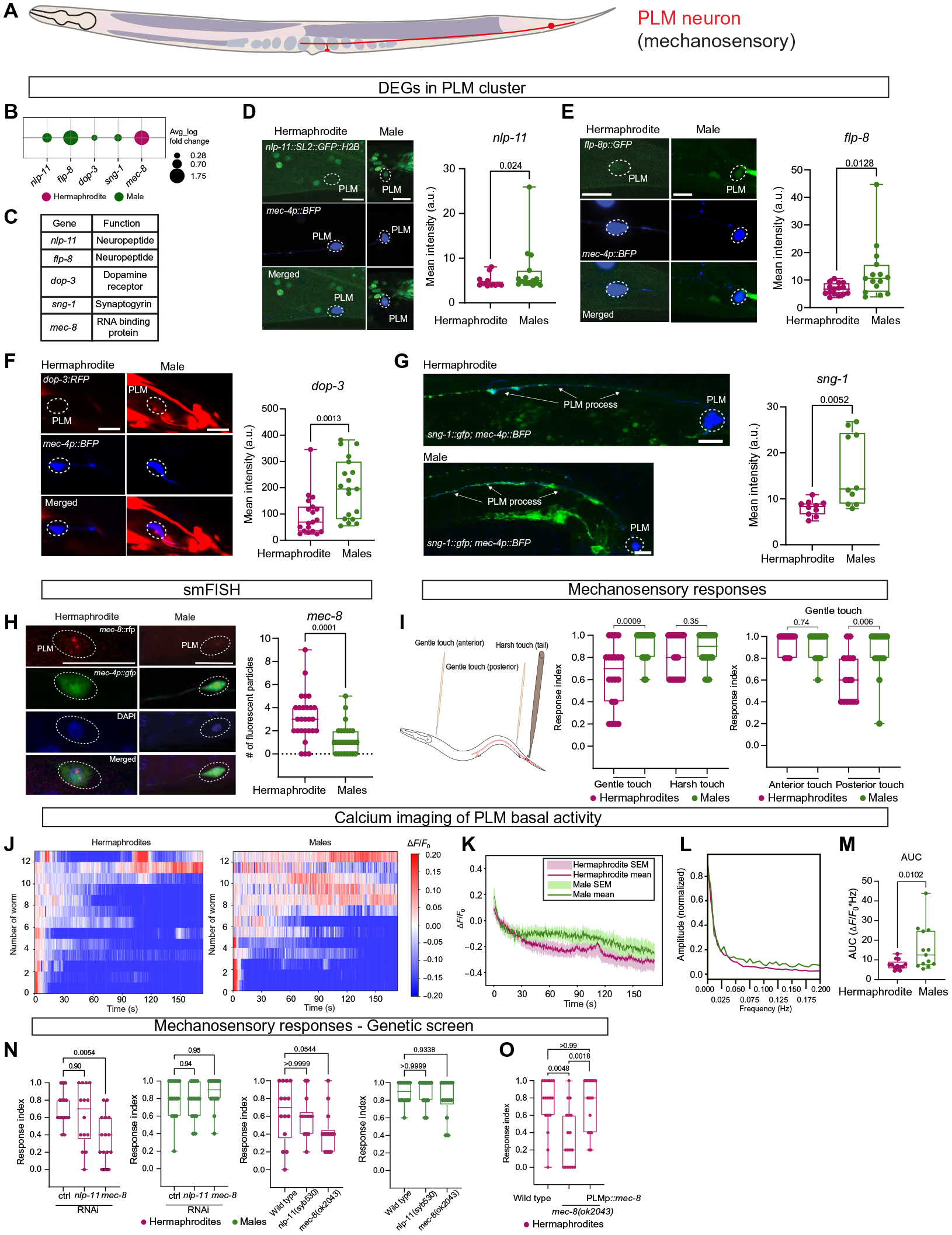
Fig. 5 PLM神经元的两性二态性。
7. 在雄性PHC神经元中,细胞粘附分子lad2是神经突正常延伸所必需的,而雌雄同体PHC神经元则不需要
接下来他们关注了PHC尾部感觉神经元,与雌雄同体相比,雄性的PHC尾部感觉神经元的神经突延伸长度是其两倍。这种伸长可能是为了适应雄性特异性神经元突触连接的增加(图4D和6A)。TF DMD-3 (DM结构域家族-3)被证明可以改变雄性PHC神经元的功能和分子特征,而在雄性PHC簇中,dmd-3、eat-4/VGLUT和神经肽flip -11的表达也显著增加(图6B)。通过排序分析,他们确定了209个DEGs(13个雄性神经元和16个雌雄同体神经元;图2A和表S2)。这些DEGs在GO术语“突触功能”和“跨膜运输”中富集(图6C)。对于scRNA-seq预测的神经元DEGs子集,包括神经肽基因,γ-氨基丁酸(GABA)和多巴胺受体,以及细胞粘附分子lad2,也进行了验证实验(图6,D至I)。
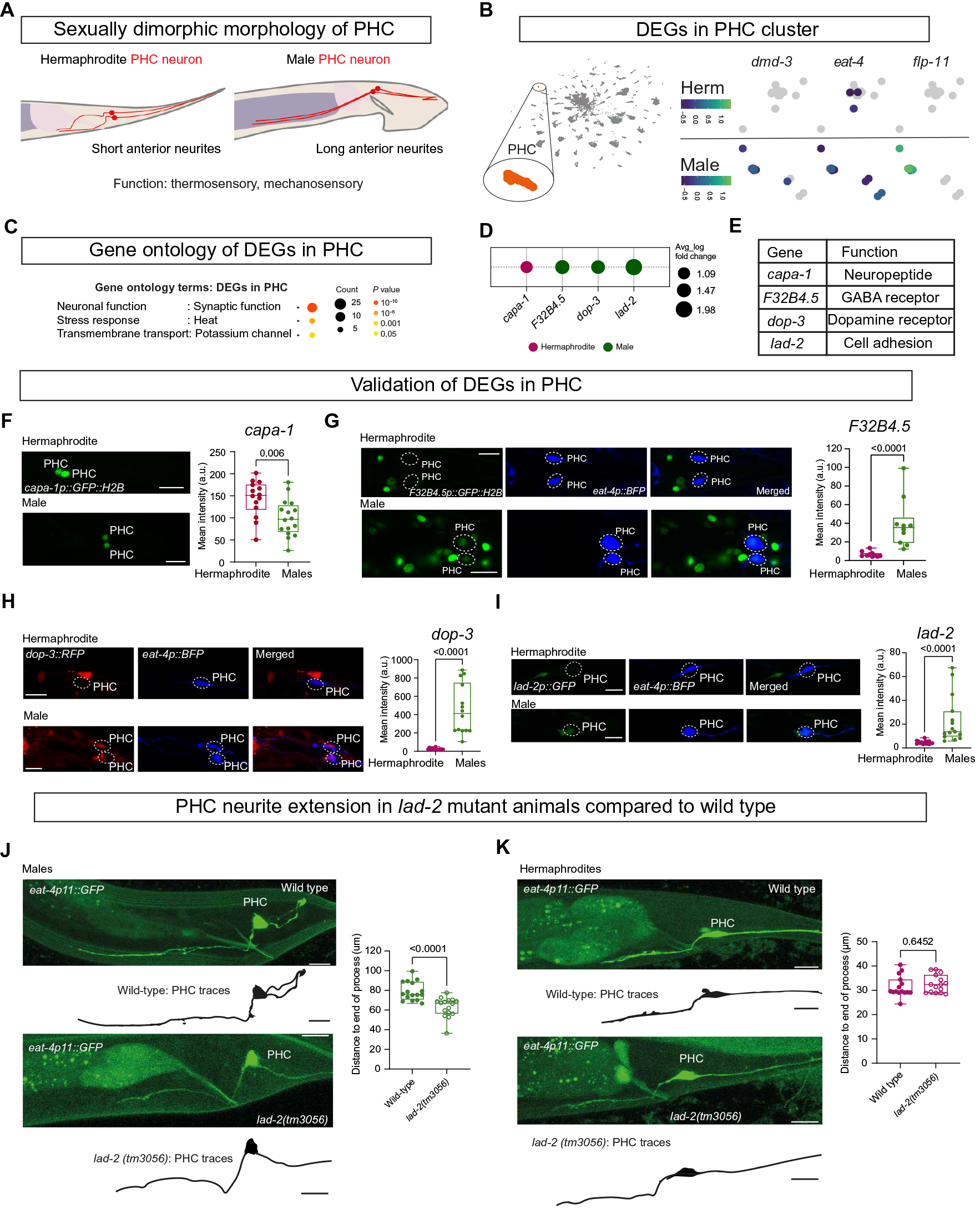
Fig. 6 DEGs在PHC中的表达。
总之,该数据集提供了一个分子框架,可以用于理解细微的基因表达及调节差异是如何调节保守的信号通路以驱动机体的性别特异性行为的。
 技术咨询:
技术咨询: